- Service Details
- Case Study
What Is Glycolysis Pathway
Glycolysis pathway is a common metabolic pathway in both prokaryotic and eukaryotic cells. It takes place in the cytosol in eukaryotic cells. By converting glucose into two molecule of the three‐carbon compound pyruvate and generating two ATPs, glycolysis pathway generates the ATP needed for energy‐requiring processes and the pyruvate needed for the complete oxidation of glucose to carbon dioxide and water. For cells lack of mitochondria such as red blood cells, the glycolysis is the only energy source of ATP. For cells containing mitochondria, glycolysis serves as a preparatory pathway for the complete oxidation of glucose to generate larger amounts of ATP. This pathway often serves as a main energy source for cancer cell. Hence, glycolysis pathway is being extensively studied in cancer research, by analyzing the glycolytic enzymes expression level and monitoring the amount of metabolites.
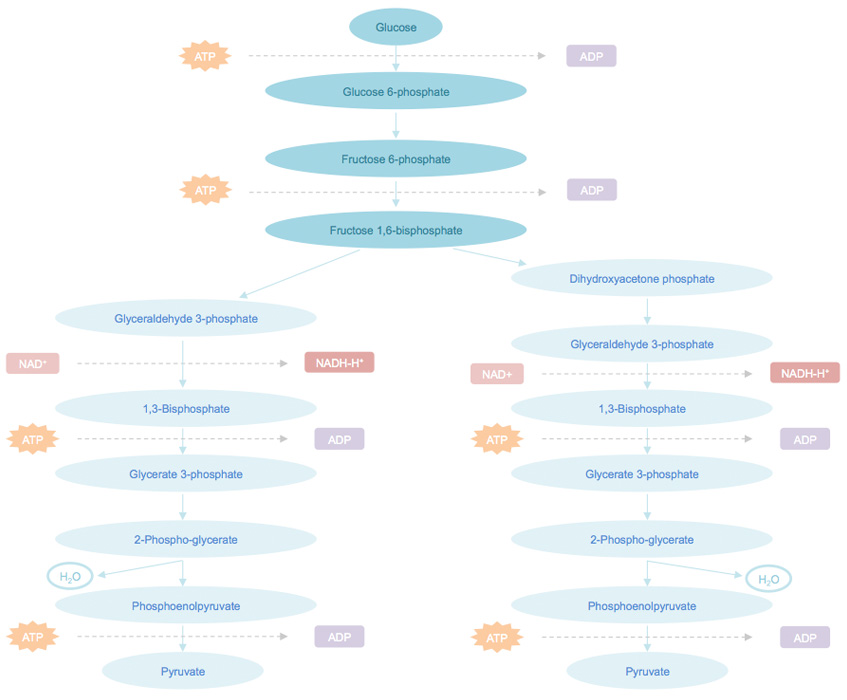
Glycolysis pathway can be divided into three stages. Stage 1, phosphorylated by ATP, glucose converts to glucose 6-phosphate, and then to fructose 1,6-bisphosphate. In this stage, after a phosphorylation, an isomerization, and a second phosphorylation reaction, glucose is trapped and converts into a compound that can be easily cut into phosphorylated three-carbon units. Stage 2, the generated fructose 1,6-bisphosphate can be cleaved into two three-carbon fragments, which are readily convertible to each other. Stage 3, ATP is generated when the phosphorylated three-carbon units are oxidized to pyruvate.
Our Glysolysis Pathway Metabolites Analysis Service
LC–MS/MS platform enable simultaneous relative and absolute quantification of multiple metabolites in this pathway. The results are widely used for investigating the relative changes in metabolite concentrations, and potential biomarker discovery. By providing the exact molecular weights and retention time, LC–MS/MS techniques serves as a powerful analytical tool for identification and quantification of metabolites. Creative Proteomics has established sensitive, reliable, and accurate HPLC-MS/MS method for quantification of glycolysis pathway metabolites.
Feature and Advantage of glysolysis pathway metabolites analysis service:
- Cutting-edge facilities
- Reliable & Reproducible
- Highly sensitive: detect low abundance metabolites
- Comprehensive analysis of metabolites
Platform
- HPLC-MS/MS
Summary
- Identification and quantification of glycolysis pathway metabolites by HPLC-MS/MS
| Glycolysis Pathway Metabolites Quantified in This Service | ||
|---|---|---|
| 1,3 phosphate glycerate/2,3 phosphate glycerate | 3 phosphate glcerate/2 phosphate glycerate | Dihydroxyacetone phosphate |
| Fructose 6 phoshpate | Fructose 1 phoshpate | Fructose 1,6 bisphosphate |
| Glucose | Glucose 6 phosphate | Glyceraldehyde 3 phosphate |
| Phosphoenol pyruvate | Pyruvate | |
Sample Requirement
| Sample |
|---|
| Animal and clinical tissue samples |
| Blood samples (serum, plasma and whole blood) |
| Urine samples |
| Stool and intestinal contents |
| Body fluid samples (cerebrospinal fluid, saliva, etc.) |
| Plant tissue samples (roots, stems, leaves flowers and fruits, etc.) |
| Cells and microbial organisms |
| Culture media and fermentation broth |
Report
- A detailed technical report will be provided at the end of the whole project, including the experiment procedure, MS/MS instrument parameters
- Analytes are reported as uM or ug/mg (tissue), and CV's are generally<10%
- The name of the analytes, abbreviation, formula, molecular weight and CAS# would also be included in the report.
With integrated set of separation, characterization, identification and quantification systems featured with excellent robustness & reproducibility, high and ultra-sensitivity, Creative Proteomics provides reliable, rapid and cost-effective glycolysis pathway metabolites targeted metabolomics services.
Impact of Up- and Downregulation of Metabolites and Mitochondrial Content on pH and Color of the Longissimus Muscle from Normal-pH and Dark-Cutting Beef
Journal: Journal of Agricultural and Food Chemistry
Published: 2020
Abstract
Sensory experience influences consumer choices, particularly evident in the sales process of fresh beef, where color deviation can lead to discounted value. Due to inherent biochemical variations, fresh beef does not always exhibit a bright red color. Black-cut beef is one example of color variation in meat, where beef does not display the typical fresh red color. Black-cut beef is produced worldwide and causes economic losses in the animal and food industries. The mechanism behind its formation remains unclear, although it is widely believed that chronic pre-slaughter stress in animals reduces glycogen reserves, resulting in an increase in postmortem muscle pH. Additionally, mitochondria in the muscles of slaughtered animals remain active, which may affect the oxidation-reduction state of myoglobin, leading to the formation of dark red deoxymyoglobin. This study aims to elucidate the biochemical differences between normal pH beef and black-cut beef by examining the differences in metabolite distribution and mitochondrial content.
Results
Through metabolite profiling, the authors discovered that compared to normal pH beef, black-cut beef exhibited downregulation of glycolytic metabolites (such as glucose-1-phosphate and glucose-6-phosphate) and upregulation of tricarboxylic acid substrates (such as malic acid and fumaric acid). Immunological analysis indicated that black-cut beef had a higher proportion of oxidative fiber types compared to normal pH beef. Moreover, black-cut beef showed higher levels of mitochondrial proteins and DNA. The increase in mitochondrial content may, to some extent, influence the oxygen consumption, myoglobin oxidation, and external appearance of dark-cut beef. The study results demonstrate that alterations in tricarboxylic acid metabolites and mitochondrial content in black-cut beef can impact muscle pH and color.
Biochemical Characteristics of Normal pH and Black-cut Beef
In this study, black-cut beef exhibited a lower L* value (L value represents lightness, with lower values indicating darker color) compared to previous reports (Table 1). The increase in pH value of black-cut beef, compared to normal pH beef, can enhance oxygen consumption and myoglobin reduction activity. In exercise physiology, mitochondrial biogenesis is an adaptive mechanism in response to chronic stress and energy deprivation. Therefore, animals exposed to stress before slaughter may increase their mitochondrial content as an organismal adjustment. Hence, one of the objectives of this study is to compare the mitochondrial content between normal pH and black-cut beef using qPCR and differential centrifugation.
Table 1: Comparison of the longest muscle mitochondrial content and biochemical characteristics between normal pH and black-cut beef.
| Parameter | Normal pH Beef | Black-cut Beef | SEM | P-value |
|---|---|---|---|---|
| (a) Composition Analysis | ||||
| Moisture Content (%) | 66.2 | 71.2 | 0.9 | <0.001 |
| Protein (%) | 22.2 | 21.6 | 0.4 | 0.51 |
| Fat (%) | 7.9 | 7.3 | 0.5 | 0.21 |
| (b) Mitochondria | ||||
| Mitochondrial Protein (mg/g tissue) | 1.85 | 2.62 | 0.13 | 0.03 |
| Mitochondrial DNA Relative Content | 1 | 2.04 | 0.48 | 0.02 |
| (c) Surface Color | ||||
| Brightness (L) | 40.5 | 29.4 | 0.84 | <0.001 |
| Redness | 28.4 | 21.5 | 0.93 | <0.001 |
| Yellowness | 22.7 | 17.1 | 0.74 | <0.001 |
| Chroma Concentration | 37.6 | 29.1 | 0.92 | <0.001 |
| Hue | 36.8 | 35.6 | 0.33 | 0.01 |
| (d) Biochemical Characteristics | ||||
| pH | 5.6 | 6.3 | 0.06 | <0.001 |
| Myoglobin Concentration (mg/g meat) | 5.7 | 7.9 | 0.27 | <0.001 |
| Oxygen Consumption (OC) | 0.82 | 1.42 | 0.2 | <0.001 |
| Myoglobin Reduction Activity (MRA) | 1.2 | 0.8 | 0.11 | <0.001 |
Differences in Metabolite Profiles between Normal pH and Black-cut Beef
The authors compared the metabolite profiles between black-cut beef and normal pH beef to assess key metabolites contributing to the color difference (Figure 1). Previous studies have shown that the occurrence of black-cut beef is closely related to high pH (>5.8) in the muscles due to the depletion of muscle glycogen pre-slaughter. In this study, black-cut beef exhibited downregulation of metabolites in the glycolytic pathway, including glucose-6-phosphate, glycerol-3-phosphorylcholine, gluconic acid, glucose, fructose, glucose-1-phosphate, fructose-6-phosphate, and galactose-6-phosphate. The downregulation of these metabolites indicates a reduced synthesis of metabolites in the glycolytic pathway, leading to a decrease in lactate production. The anaerobic environment in the muscle favors lactate accumulation, resulting in a decrease in post-slaughter pH. Muscles with high pH are associated with lower levels of glucose-6-phosphate and lower anaerobic glycolytic rates. Therefore, the downregulation of glycolytic flux in the muscle limits its ability to produce lactate. Consequently, the lower levels of glycolytic metabolites can explain the observed high pH in black-cut beef.

The depletion of metabolites in the glycolytic pathway promotes the conversion of alternative sources into the glycolytic pathway to supply energy to the organism. In this study, compared to normal pH beef, black-cut beef exhibited excessive levels of pyruvic acid. Additionally, the authors observed upregulation of metabolites in the TCA (tricarboxylic acid) cycle in black-cut beef, including fumaric acid, malic acid, and citric acid. Fumaric acid is converted to malic acid, which is further converted to oxaloacetic acid, generating NADH. The upregulation of TCA cycle metabolites indicates that black-cut beef has a stronger ability to generate ATP through the utilization of metabolites.
In this study, upregulation of the ATP metabolite xanthine was observed in black-cut beef, indicating better energy utilization compared to normal pH beef. Nelson et al. suggested that under stress conditions, energy stores are depleted due to endocrine processes such as hormone secretion. Consequently, cellular adaptive adjustments promote mitochondrial biogenesis and related pathways to restore cellular energy levels. Therefore, the observed downregulation of energy storage-related metabolites, upregulation of energy synthesis metabolites, and higher levels of mitochondrial DNA and proteins in black-cut beef can be attributed to these adaptations. However, the activation and regulation of mitochondrial bioenergetics and biogenesis after animal slaughter remain unclear. Other metabolites, including gamma-aminobutyric acid (GABA), succinic semialdehyde, and nucleotide metabolites (ribose-5-phosphate, uracil, xanthine, and cytidine), were upregulated in black-cut beef. GABA acts as an inhibitory neurotransmitter and counteracts glutamate-mediated excitotoxicity by opening calcium channels. Succinic semialdehyde is a GABA metabolite formed by transaminase action and can be further oxidized to succinic acid, entering the TCA cycle for energy synthesis. The upregulation of succinic semialdehyde in black-cut beef suggests a possible deficiency of succinic semialdehyde dehydrogenase (SSADH). SSADH deficiency is associated with reduced succinic acid synthesis. Therefore, a downregulation of succinic acid metabolites is observed in black-cut beef. Additionally, SSADH deficiency is associated with a rare autosomal recessive genetic disorder in the GABA degradation pathway, involving cerebellar ataxia, a neurological symptom characterized by the lack of voluntary muscle coordination. Therefore, the authors speculate that black-cut beef may have higher levels of involuntary coordinated muscle movements, which could consume energy storage-related metabolites and increase the demand for energy synthesis-related metabolites.
Conclusion
In conclusion, black-cut beef exhibits higher mitochondrial content and downregulation of metabolites such as glucose-6-phosphate, fructose-6-phosphate, and glucose-1-phosphate in the glycolytic pathway. The decreased synthesis of lactate during anaerobic metabolism contributes to the elevated pH in black-cut beef. Additionally, TCA metabolites such as fumaric acid, citric acid, and malic acid are upregulated, which can activate mitochondrial activity and limit the binding of myoglobin to oxygen.







